- Genetic resources of teak (Tectona grandis Linn. f.)—strong genetic structure among natural populations. Centre of diversity in semi-moist eastern coast of India and in Myanmar.
- Analysis of Genetic Diversity and Population Structure of Rice Germplasm from North-Eastern Region of India and Development of a Core Germplasm Set. 10% of 7000 very diverse accessions recovers 99.9% of allelic diversity.
- Global assessment of urban and peri-urban agriculture: irrigated and rainfed croplands. Urban croplands represent 6% of total. No word on what percentage of genetic diversity, but I bet more than 6%.
- Satellite Earth observation data to identify anthropogenic pressures in selected protected areas. Some data can be used to identify some pressures.
- Global diversity and geography of soil fungi. Climatic factors are key, but look at the diagram.
- Biosynthesis, regulation, and domestication of bitterness in cucumber. So that Dutch breeder, who Jeremy always mentions, who once tasted 10,000 cucumber cotyledons in his quest to breed a nice-tasting one, would not have to do so now. Which is a pity.
Nibbles: Chocolate, MAS, Cash crops, Medicinal plants, Rice domestication, Cat genome, Banana research, Artichoke history, Root vegetables, Diabetes data, eMonocot, Paris herbarium, Appleseed, Seed saving, Potato safety duplication, Seed atlas, Botanical Jurassic Park, Mapping urban fruit, Midwest road trip, Iraqi marshes, World Digital Library, World Parks Congress, Plant demography
- Ok here goes, there’s a week’s work of Nibbles we’ve got to catch up on.
- World running out of chocolate! Tell that to Cologne.
- Yeah well I prefer tea to cocoa.
- Greenpeace: “Smart breeding” will save us, not GMOs. Breeders: All breeding is smart.
- Guess the world’s biggest cash crop. Yeah, that one.
- Alas, it’s not included in the recent strategy for conserving medicinal plants. Not that it would need conserving.
- The domestication of the world’s biggest crop, period. Deconstructed. And if you want to drill down.
- And of the world’s biggest fruit.
- And of the world’s biggest pet.
- And of the world’s most difficult to eat vegetable.
- Root “vegetables” made simple. Because winter.
- And why you must eat your veggies, including the difficult ones.
- Videos on Kew’s monocot database and on the renovation of another famous herbarium.
- The real Johnny Appleseed.
- But you too can save seeds, just like Johnny.
- But don’t forget to safety duplicate, like CIP has done, at Embrapa.
- And this shows you what those seeds can look like.
- You don’t necessarily need seeds to save plants, though.
- Mapping fallen fruit. Because we can.
- Road trip!
- Boat trip!
- Selected Techniques in the Art of Agriculture. From French to Turkish to Arabic. One of many nifty agriculture-related resources in the World Digital Library.
- Oh yeah, the IUCN World Parks Congress has been on and its all over the intertubes. Including with neat visualizations, natch.
- How many of the species in the COMPADRE database of plant demography information are in protected areas? How many are crop wild relatives? I need an intern.
Brainfood: Enset & cattle, Evolution Canyon, Indian spices, Bohemian fruit rhapsody, ILRI forage genebank, Wild sunflower, Agroecology, Holistic hazelnuts, Culture & conservation, Salty broomcorn, Fancy mapping, German cherries, Ethiopian barley nutrients
- Sidama Agro-Pastoralism and Ethnobiological Classification of its Primary Plant, Enset (Ensete ventricosum). The Sidama feed the high-protein parts of enset to cattle and then get their protein from milk. Seems a roundabout way of going about things but I guess they know best.
- Evolution of wild barley at “Evolution Canyon”: adaptation, speciation, pre-agricultural collection, and barley improvement. One-stop shop for researching evolution of a crop wild relative.
- Spices and Condiments: Status of Genetic Resources and Setting Priorities for Introduction in India. National Bureau of Plant Genetic Resources is on the job, collecting at home and acquiring from genebanks abroad.
- Inventory and conservation of fruit tree landraces as cultural heritage of Bohemian Forest (Czech Republic), indicators for former settlements of ethnic minorities. That would mean Germans. No word on whether the database has been cross-checked with that of BLE-IBV. Interested in the topic of European landraces in general? Try this from Bioversity.
- Forage Diversity: An Essential Resource to Support Forage Development. ILRI’s genebank deconstructed.
- Wild Sunflower Species as a Genetic Resource for Resistance to Sunflower Broomrape (Orobanche cumana Wallr.). Pretty much all the perennial species have resistance, and many of the annuals. Thank goodness for the USDA collection, eh?
- Agroecological Research: Conforming — or Transforming the Dominant Agro-Food Regime? Bit of both? Is that such a bad thing?
- A multidisciplinary approach to enhance the conservation and use of hazelnut Corylus avellana L. genetic resources. Holistic, even.
- The Cooked is the Kept: Factors Shaping the Maintenance of Agro-biodiversity in the Andes. Keep your culture, keep your crop diversity.
- Response of broomcorn millet (Panicum miliaceum L.) genotypes from semi arid regions of China to salt stress. 39 out of a core collection of 195. Result!
- Vital Signs: Integrating Data To Visualize the Human, Agriculture, and Nature Nexus. Sounds promising enough an effort to bring together livelihoods, production and environmental data, but when you go to the website (for Tanzania in this case), all you get is a bunch of admittedly very pretty pdf maps.
- Phenotypic and genotypic characterization in the collection of sour and duke cherries (Prunus cerasus and ×P. ×gondouini) of the Fruit Genebank in Dresden-Pillnitz, Germany. …give different results. If I had a dollar…
- Genome-wide association mapping of zinc and iron concentration in barley landraces from Ethiopia and Eritrea. There are QTLs. Now what?
Nibbles: Cannabis, Brachiaria, Grasslands, Oryza, Taxonomy resources, Artocarpus, Quercus, Zea, MAS, GBIF
- “Something researchers are looking at is which cultivars, or strains, of hemp are best for the various uses — fiber, oil, nutrition, etc.” Love that etc.
- Speaking about grass: Brachiaria goes home, to wild acclaim.
- Did someone say wild? Wild grass needs help!
- Rice is a grass. Oh my yes.
- How to keep up to date with taxonomic research online.
- Pacific Regional Breadfruit Initiative gets an award.
- You can also make flour from acorns.
- And maize: what’s a grit?
- Greenpeace touts MAS.
- Next thing you know they’ll be singing the praises of Big Data. Yeah, maybe not.
The identity of “millet” in East Africa
A blog post from the Chicago Council on Global Affairs a couple of days ago reminded me that we had blogged about GYGA — the Global Yield Gap and Water Productivity Atlas — a couple of years back. Just to remind us all
…the target of the Global Yield Gap Atlas (GYGA) is to provide best available estimates of the exploitable yield gap (Yg-E) — difference between current average farm yields and 80% of Yp and Yw. Water resources to support rainfed and irrigated agriculture also are limited, which means efficiency in converting water to food, water productivity (WP), is another key food security benchmark included in the Atlas.
I’ve had a quick look at the GYGA website, and it seems serviceable enough, though it has the drawbacks, often remarked on in these pages, that it is difficult to share the maps you make, and import your own data into them. Here I’d like to point out another potential issue, though. This is what you get when you look at the yield gap for “rainfed millet.” Because of the aforementioned drawbacks, a clunky screenshot is the best I can manage, I’m afraid.
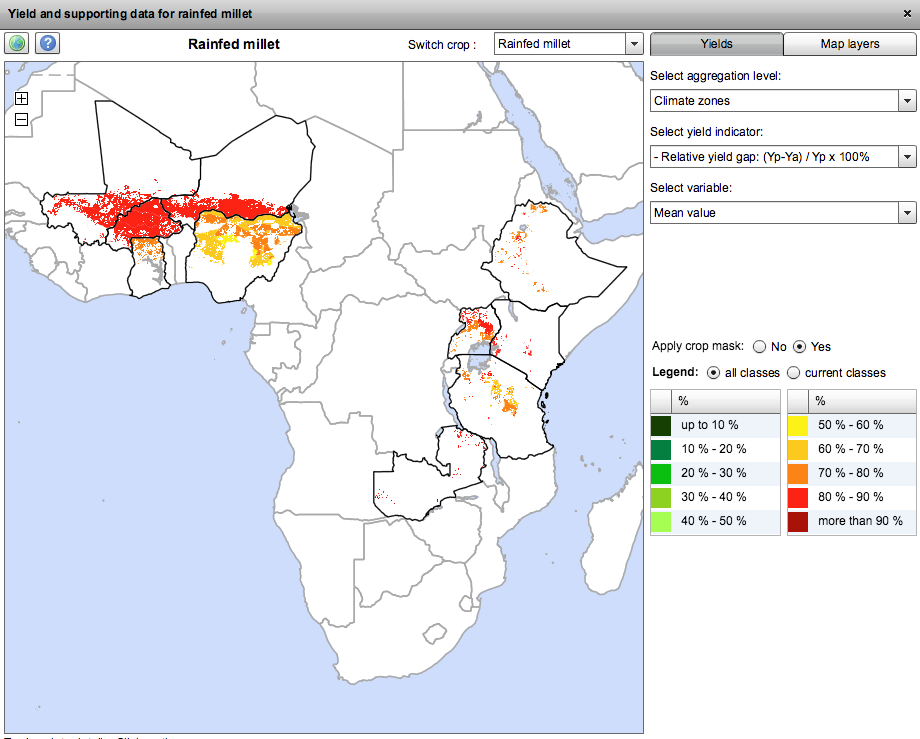
Now, the stuff in West Africa is clearly pearl millet, but what about in East Africa? Is there really that much pearl millet in Uganda, say? This is what Genesys knows about the two millets in East Africa. And yes, it’s a clunky screen grab too, but if I had wanted to, I could have downloaded separate KML files for the two crops and opened them both in Google Earth and then exported a nice JPG.
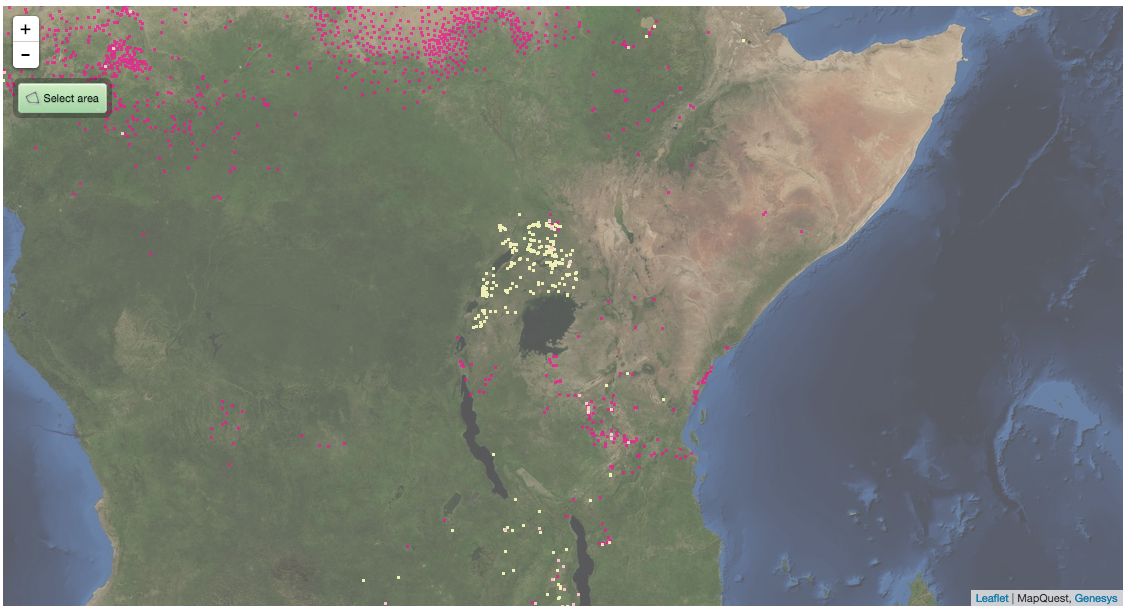
Pink is pearl, yellow is finger. There’s a little bit of pearl millet in Uganda, but not all that much. The millet there is mainly finger millet. So, which is the millet in GYGA? Is it confusing the two?