- On a day that we hope will be historic for the right reason…
- The history of the sweet potato.
- The history of the horse.
- The history of the study of plant diversity. And more.
- The history of IR8.
- The banana is not yet history.
- The history of vegetables. And more.
- The continuing history of Iroquois corn.
Brainfood: Truffle diversity, Pig diversity, SDMs
- How the truffle got its mate: insights from genetic structure in spontaneous and planted Mediterranean populations of Tuber melanosporum. More outbreeding in plantations, but strong structure still.
- Comparing genetic diversity of pig populations on the US mainland, Pacific Islands and China: Y chromosome evaluation. 16 breeds, 5 haplotypes, 10 breeds with only 1.
- What we use is not what we know: environmental predictors in plant distribution models. Edaphic factors are being ignored. ‘Twas ever thus.
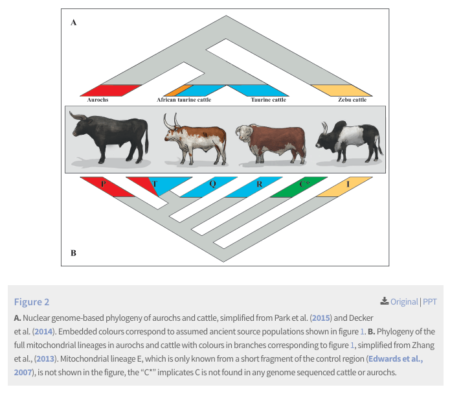
Aurochs redux
A paper just out in Open Quaternary discusses “The Draft Genome of Extinct European Aurochs and its Implications for De-Extinction.” Not to boast about it, but we were onto that five years back.
Brainfood: SE Asia archaeobotany, Avocado cryo, Farm diversity & revenue, DOC cheese, Kenyan agrobiodiversity, Perennial pigeonpea, Algerian sheep diversity, Basil rankings, Wild sunflower
- Rice, beans and trade crops on the early maritime Silk Route in Southeast Asia. At trade crossroads, crops had to audition for inclusion in the local menu.
- Cryopreservation of somatic embryos for avocado germplasm conservation. Still needs work.
- Soil fertility, crop biodiversity, and farmers’ revenues: Evidence from Italy. Diverse farms are more profitable, and can make up for poor soils.
- Native and Non-Native Sheep Breed Differences in Canestrato Pugliese Cheese Quality: a Resource for a Sustainable Pastoral System. Traditional local cheese is better when made with milk from traditional local breeds.
- Agrobiodiversity conservation enhances food security in subsistence-based farming systems of Eastern Kenya. But correlation is not causation.
- Estimating demand for perennial pigeon pea in Malawi using choice experiments. It won’t be liked everywhere.
- Genome-wide analysis highlights genetic dilution in Algerian sheep. Two of the 7 local breeds studied are in trouble due to uncontrolled breeding with a third, but may be doing better in neighbouring countries.
- Multicriteria optimization to evaluate the performance of Ocimum basilicum L. varieties. Fancy maths allows you to pick the best basil variety out of 8.
- The challenges of maintaining a collection of wild sunflower (Helianthus) species. Are many and varied…
Nibbles: European bison, Eucalypts and CC, Flood prevention, Svalbard, Cute animals
- What sort of sick bastard decapitates European bison re-introduced to the wild?
- Might have to introduce eucalypts to different places soon. No doubt that will also end in tears.
- Diversity helps with flooding. But it’s probably too late.
- USDA buys some insurance. Because life.
- Animal portraits are guaranteed to make your day better. Even this day.